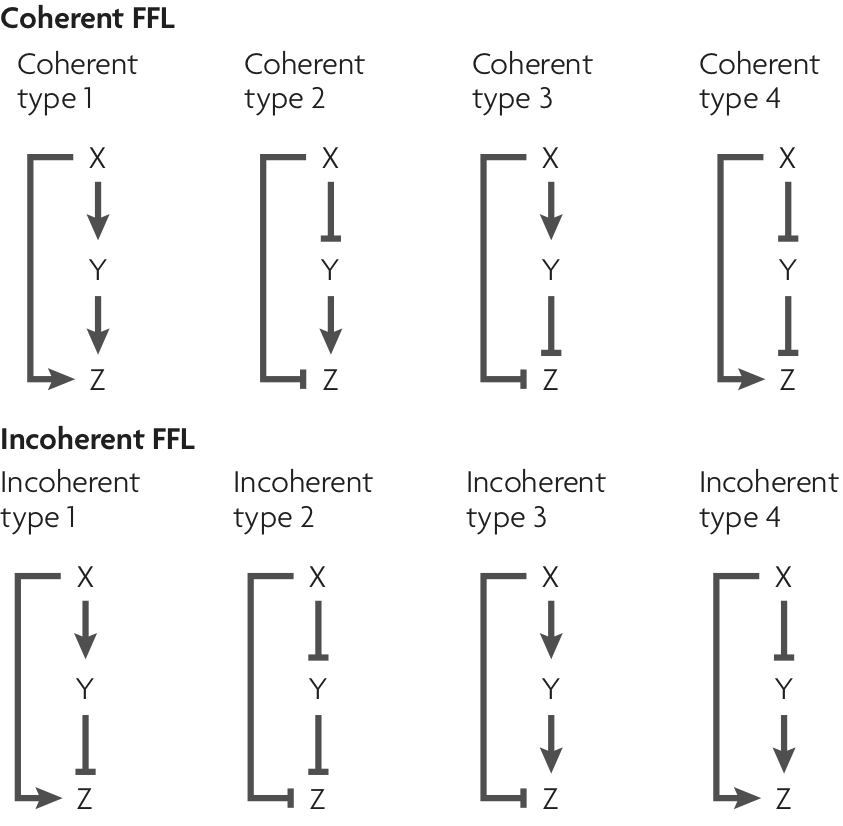
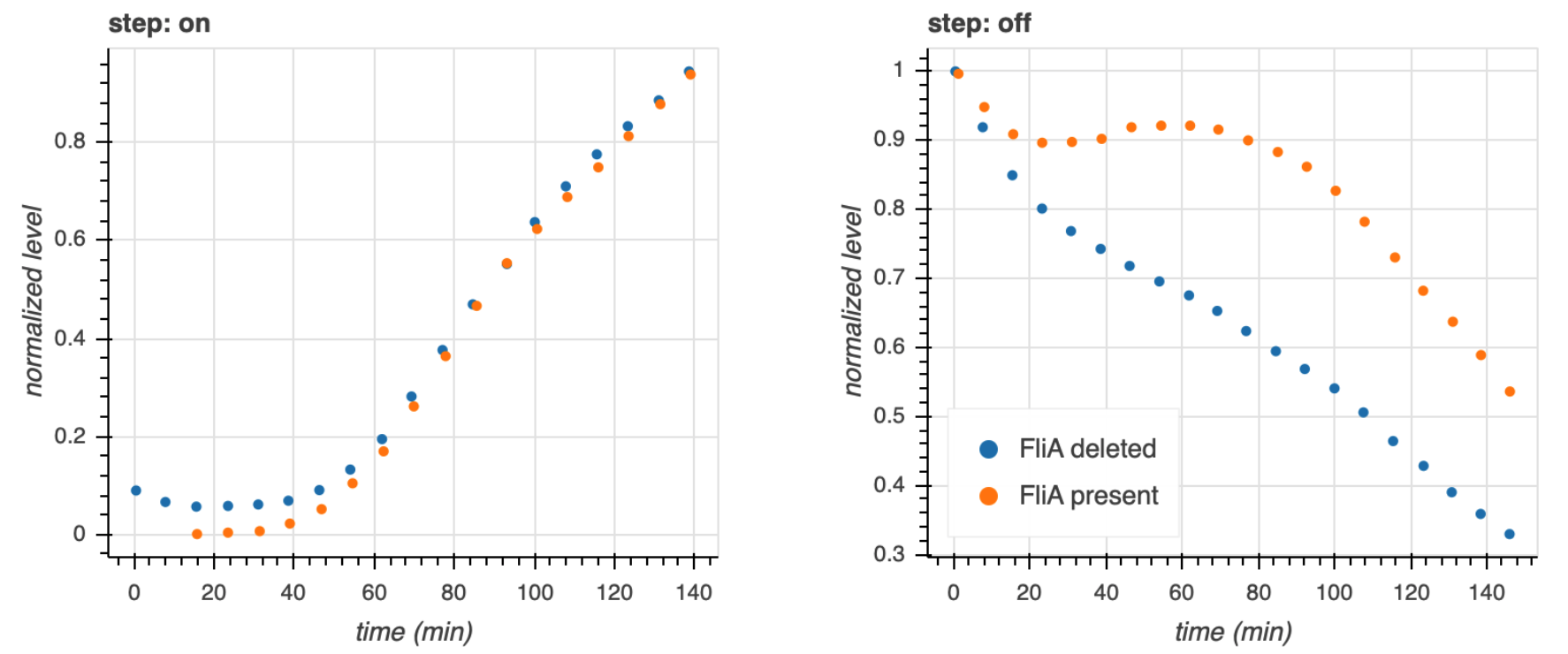
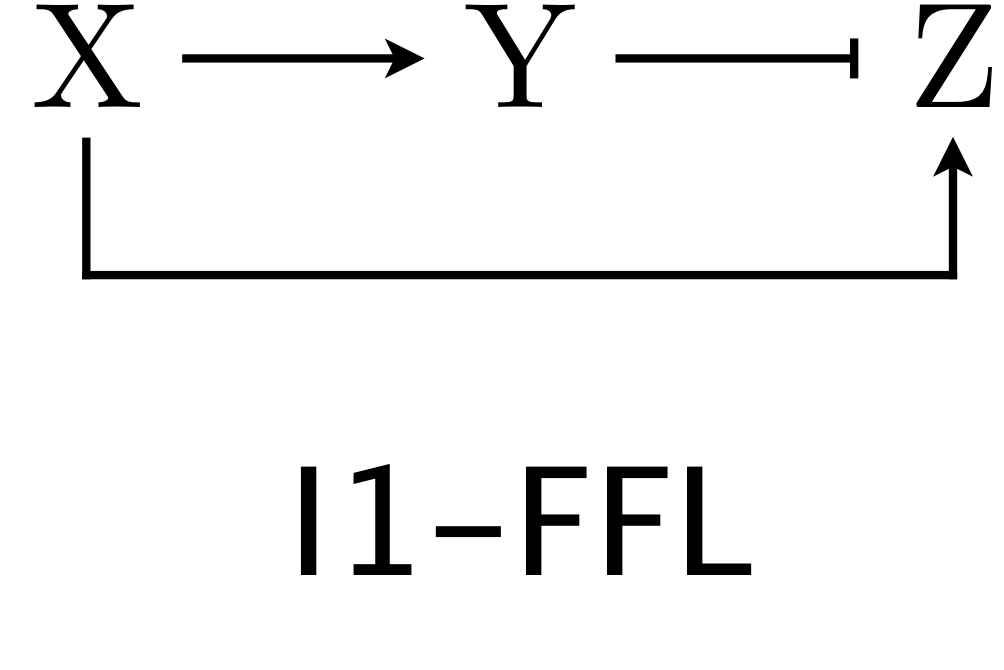
Dynamical equations for FFLs¶
To analyze the C1-FFL and I1-FFL (or any of the other FFLs), in response to changes in the input X, we can write a generic system of ODEs for the concentrations of Y and Z. We know that Y is either activated or repressed by X and itself experiences degradation. We define a dimensionless function $f_y(x/k_{xy}; n_{xy})$ to describe the activating or repressive Hill function for the regulation of X by Y. We have used the notation that $n_{ij}$ is the Hill coefficient for j regulated by i, with $k_{ij}$ similarly defined. To be explicit:
If X activates Y, then
\begin{align}
f_y(x/k_{xy}; n_{xy}) = \frac{(x/k_{xy})^{n_{xy}}}{1 + (x/k_{xy})^{n_{xy}}}\enspace.
\end{align}
If X represses Y, then
\begin{align}
f_y(x/k_{xy}; n_{xy}) = \frac{1}{1 + (x/k_{xy})^{n_{xy}}}\enspace.
\end{align}
The dynamical equation for $y$ for either activating or repressive action by X is
\begin{align}
\frac{\mathrm{d}y}{\mathrm{d}t} &= \beta_y\,f_y(x/k_{xy}; n_{xy}) - \gamma_y y.
\end{align}
Similarly, we define the dimensionless function $f_z(x/k_{xz}, y/k_{yz}; n_{xz}, n_{yz})$ to describe the regulation in expression of Z by X and Y. This have any of the functional forms we have listed above for activation/repression pairs and AND/OR logic. The dynamical equation for $z$ is then
\begin{align}
\frac{\mathrm{d}z}{\mathrm{d}t} &= \beta_z\,f_z(x/k_{xz}, y/k_{yz}; n_{xz}, n_{yz}) - \gamma_z z.
\end{align}
We can nondimensionalize these equations by choosing
\begin{align}
t &= \tilde{t} / \gamma_y, \\[1em]
x &= k_{xz}\,\tilde{x},\\[1em]
y &= k_{yz}\,\tilde{y},\\[1em]
z &=z_0\,\tilde{z},
\end{align}
where $z_0$ is as of yet unspecified. Inserting these expressions into the dynamical equations gives
\begin{align}
\gamma_y k_{yz}\,\frac{\mathrm{d}\tilde{y}}{\mathrm{d}\tilde{t}} &= \beta_y\,f_y\left(\frac{k_{xz}}{k_{xy}}\,\tilde{x}; n_{xy}\right) - \gamma_y k_{yz} \tilde{y},\\[1em]
\gamma_y z_0\,\frac{\mathrm{d}\tilde{z}}{\mathrm{d}\tilde{t}} &= \beta_z\,f_z(\tilde{x}, \tilde{y}; n_{xz}, n_{yz}) - \gamma_z z_0 \tilde{z}.
\end{align}
If we conveniently define $z_0 = \beta_z/\gamma_z$, then the dynamical equations become
\begin{align}
\frac{\mathrm{d}\tilde{y}}{\mathrm{d}\tilde{t}} &= \beta\,f_y\left(\kappa\tilde{x}; n_{xy}\right) - \tilde{y},\\[1em]
\gamma^{-1}\,\frac{\mathrm{d}\tilde{z}}{\mathrm{d}\tilde{t}} &= f_z(\tilde{x}, \tilde{y}; n_{xz}, n_{yz}) - \tilde{z},
\end{align}
where we have defined
\begin{align}
&\beta = \frac{\beta_y}{\gamma_y k_{yz}},\\[1em]
&\gamma = \frac{\gamma_z}{\gamma_y}, \\[1em]
&\kappa = \frac{k_{xz}}{k_{xy}}.
\end{align}
In addition to the Hill coefficients, these dimensionless parameters complete the parameter set of the FFL dynamical system. Each has a physical meaning. The parameter $\beta$ is the dimensionless unregulated steady state level of $y$, $\gamma$ is the ratio of the decay rates of Z and Y, and $\kappa$ is the ratio of the amounts of X that are necessary to regulate Z and Y.
Henceforth, we will work with these dimensionless equation and will drop the tildes for notational convenience.